












































































































Si è già affermato che è grazie alla fotosintesi clorofilliana che l’energia solare viene intrappolata entro gli ecosistemi, e che è grazie ad essa che tutti gli organismi di un ambiente possono vivere e riprodursi; è giunto ora il momento di vedere come si verifica in realtà questo fenomeno.
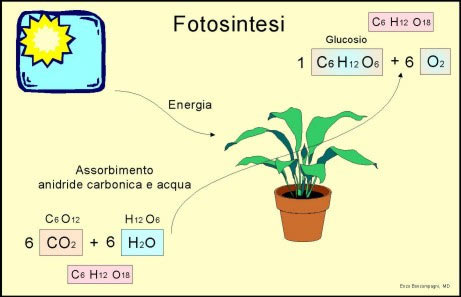
Pigmenti fotosinteticiLa fotosintesi si verifica nei cloroplasti, il cui colore dipende dalla pre-senza della clorofilla, che è la molecola chiave di tutto il processo.La clorofilla è costituita da quattro anelli tetrapirrolici uniti fra loro; nel centro è presente un atomo di magnesio. Ad uno degli anelli è associata una molecola di fitolo, un idrocarburo a lunga catena che ha lo scopo di ancorare la clorofilla alla membrana interna dei cloroplasti. Esistono vari tipi di clorofilla, che differiscono l’una dall’altra per la natura di alcuni gruppi legati in una parte dell’anello; è in base al tipo di clorofilla posseduta che gli esseri autotrofi vengono classificati in gruppi omogenei ed affini dal punto di vista evolutivo. Oltre alla clorofilla nel processo fotosintetico sono impegnati anche altri pigmenti, quali i carotenoidi, di colore giallo, arancio o rosso, le xantofille, gialle, e le ficobiline, blu e rosse, PROVA 12.2. Tutti questi pigmenti svolgono una funzione accessoria e di aiuto nei confronti della clorofilla, e molto spesso sono così numerosi da mascherare il tipico colore verde della clorofilla. I vari pigmenti si associano in complessi multimolecolari formati da al-meno 3007 molecole di vario tipo, che hanno lo scopo di assorbire la maggior quantità possibile di luce solare, e di trasferirla ad un’unica molecola di clorofilla a, detta centro di reazione. Solo la clorofilla a, è infatti in grado di utilizzare l’energia correttamente. I complessi di pigmenti si trovano immersi nello spessore della membrana dei filacoidi in corrispondenza dei grana, ed ad essi sono associate delle mole-cole di varia natura che costituiscono una catena fotosintetica di trasporto degli elettroni. A questo insieme viene dato il nome di fotosistema, ed è stato evidenziato come nelle piante superiori siano presenti due fotosistemi, denominati fotosistema I e fotosistema II. I due fotosistemi differiscono per il tipo di clorofilla presente nel centro di reazione: il fotosistema I possiede infatti una molecola di clorofilla detta P700, in quanto assorbe prevalentemente la luce di 700 nm di lunghezza d’onda, mentre il fotosistema Il è caratterizzato dalla clorofilla P680, che assorbe prevalentemente alla lunghezza d’onda di 680 nm. In questo modo i due fotosistemi non competono fra loro per l’assorbimento della luce solare. Utilizzando particolari tecniche di osservazione al microscopio elettronico, nella membrana dei tilacoidi è stata evidenziata la presenza di diverse migliaia di tali complessi: la superficie della membrana appare in-fatti coperta da una fitta granulazione corrispondente ai singoli fotosistemi. Fase luminosaLa luce viene considerata un’onda elettromagnetica che si propaga nello spazio trascinandosi dietro una certa quantità di energia. Le onde lumi-nose si differenziano in base sia alla loro lunghezza d’onda, espressa in nm, sia al loro contenuto energetico che risulta inversamente proprozionale ad essa: tanto maggiore è la lunghezza d’onda, tanto minore sarà la quantità di energia che l’onda è in grado di trasmettere. Quando un’onda luminosa colpisce un corpo gli cede l’energia che essa trasporta; tale energia viene assorbita dalla struttura molecolare del corpo, che subirà quindi delle modificazioni più o meno evidenti. La quantità di energia associata ad un’onda viene normalmente definita fotone e sta ad indicare, da una parte, che essa ha un valore costante, di-pendente dalla lunghezza dell’onda, e dall’altra, che l’onda stessa può trasmettere solo quantità ben definite di energia. Il fotone viene solita-mente ad interagire con gli elettroni impegnati nella formazione dei doppi legami presenti nelle molecole dei pigmenti; questi elettroni, infatti, sono trattenuti più debolmente e possono più facilmente quindi essere eccitati e passare ad un livello superiore. I vari pigmenti presenti in ciascun fotosistema assorbono tutte le lunghezze d’onda presenti nella luce che incide sui cloroplasti e poi trasferiscono l’energia così assorbita alla clorofilla presente nel centro di rea-zione. I pigmenti si comportano in pratica come una specie di “antenna” che serve a catturare tutte le possibili lunghezze d’onda e a convogliarle verso l’unica molecola che è in grado di trasformare tale eccitazione degli elettroni in processo chimico. Quando un fotone colpisce il fotosistema H, il primo a venir attivitato, la molecola di P680 viene a perdere un elettrone che compie un notevole salto energetico, tale da allontanarlo dalla molecola e da farlo catturare da una molecola accettrice di elettroni che si trova associata al fotosistema. Questo accettore ha un’affinità chimica talmente elevata per l’elettrone che la molecola di clorofilla non può più riaverlo indietro, e per tornare al suo stato normale deve cercare di sottrarre un elettrone ad un’altra molecola. Una volta che la clorofilla ha ceduto il suo elettrone, deve trovare una molecola con un potenziale più basso del suo. Nella cellula è l’acqua, composto molto abbondante, la molecola più adatta a questo scopo. Come esattamente avvenga la sottrazione di questi elettroni non è ancora chiaro, e vengono proposti meccanismi molto complessi, quel che interessa però è che la clorofilla riesce in qualche modo a sottrarre questi elettroni all’acqua, trasformandola secondo la reazione: Come si vede, oltre agli elettroni si forma anche una molecola di ossigeno, che non serve alla pianta e che quindi viene allontanata come pro-dotto indesiderato. I viventi hanno poi imparato ad utilizzare molto bene questo rifiuto de-la fotosintesi! È logico suppone che tale reazione possa completarsi solo quando due molecole di P680 sono state eccitate, dato che due sono gli elettroni che si liberano da una molecola di H20. La clorofilla a questo punto è pronta a riprendere le sue funzioni e ad eccitarsi nuovamente all’arrivo di altri fotoni. In definitiva i fotoni non servono ad altro che ad eccitare gli elettroni della clorofilla e a portarli verso un potenziale più negativo: in questo salto energetico avviene la trasformazione dell’energia luminosa in energia chimica. L’elettrone è ora associato con l’accettore primario, una molecola con un potenziale più negativo di molte altre a lui vicine nel fotosistema e passerà ad un’altra molecola che sarà poi costretta a cederla ad un’altra ancora e così via, in una serie continua di reazioni di riduzione e di ossidazione, fino a che l’elettrone verrà catturato dalla molecola di clorofilla P700 del fotosistema I. Durante questo passaggio dell’elettrone lungo la catena di trasporto pre-sente entro la membrana del tilacoide, ioni H+ presenti all’esterno del grana vengono pompati dentro, per poter bilanciare le cariche in movimento. In questo modo, tenendo anche conto che i protoni liberati dalla fotolisi dell’acqua sono rimasti nel comparto interno del grana, si viene creando quel gradiente chemiosmotico che è responsabile della sintesi di ATP. Infatti per ogni elettrone che fluisce entro la catena di trasporto uno ione H+ entra nello stroma del grana, e quindi per ogni molecola di acqua che viene ossidata entrano due protoni. Questi, sommandosi ai due liberati direttamente dall’acqua stessa danno quattro ioni H+, i quali fluiscono attraverso il canale dell’enzima ATPasi e permettono la sintesi di due ATP. Quando l’elettrone giunge al fotosistema I, l’arrivo di un fotone riesce a rieccitarlo al punto da trasferirlo ad un altro accettore a potenziale ancor più negativo. Anche qui l’eccitazione può avvenire solo grazie all’intervento del centro di reazione del fotosistema I e delle molecole antenna ad esso associate. L’accettore ridotto tende a passare l’elettrone ad un’altra molecola trasportatrice a potenziale maggiore, la quale a sua volta la passerà ad altre molecole, fino a che l’elettrone verrà catturato da una molecola di NADP+ (NAD-fosfato). Questo ultimo catturerà anche un protone dallo stroma, riducendosi parzialmente e bloccando l’elettrone, che non potrà più sfuggire. In definitiva l’elettrone sarà fluito lungo i due fotosistemi e lungo le due catene di trasportatori ad essi associati, per fermarsi a livello di questa molecola, il cui potenziale di ossidoriduzione è molto più basso rispetto a quello che l’elettrone possedeva in partenza. (La riduzione di una molecola di NADP+ richiede in realtà la cattura di due elettroni e di uno ione H+). Il NADPH è quindi un accumulatore di energia ed anche un comodo trasportatore. L’energia che esso possiede è in effetti solo una parte di quel-la contenuta nei due fotoni che hanno attivato le molecole di clorofilla, poiché l’altra parte è stata utilizzata per produrre una molecola di ATP. Si può affermare che l’energia luminosa è servita per portare un elettrone ad un potenziale di ossidoriduzione più basso del normale, contro la sua naturale tendenza. In questo modo si è verificato un accumulo di energia chimica potenziale che potrà poi essere sfruttata dalla cellula per le sue necessità. Riassumendo, grazie all’intervento di quattro fotoni si sono venute a formare due molecole di ATP e una di NADPH per ogni due elettroni trasportati. Siccome una molecola di acqua libera due elettroni, e dal momento che, per formare una molecola di ossigeno, necessitano due molecole di acqua, il bilancio finale del processo sarà: 8 fotoni +2 H2O+2 NADP++4 ADP+ 4P i — 4ATP+ 2NADPH+ 0,+ H+ Affinché si verifichino questi fenomeni è necessario l’intervento della luce, tale fase viene detta fase luminosa della fotosintesi, per distinguer-la dalla fase successiva, per la quale la presenza di luce non è indispensabile e che sarà detta fase oscura. Fase oscuraLa fase oscura della fotosintesi inizia quando nello stroma del cloroplasto si è accumulata una quantità sufficiente di ATP e di NADPH da per-mettere l’avvio della sintesi del glucosio, perciò inizia un po’ in ritardo rispetto alla fase luminosa, anche se poi si verifica contemporaneamente ad essa. Tale fase proseguirà fintanto che ATP e NADPH saranno disponibili, e quindi anche oltre la fine della fase luminosa. Il potere riducente accumulato nella molecola del NADPH viene ora utilizzato per ridurre chimicamente la molecola della CO,, che la pianta as-sorbe dal proprio ambiente, ossia dall’acqua per le piante acquatiche, o dall’ aria per quelle terrestri. Tale processo avviene secondo una serie complessa di reazioni chimiche che prendono il nome di ciclo di Calvin, dallo scienziato che per primo riuscì a definirne i passaggi. Come si sa la reazione globale della foto-sintesi è: energia solare 6CO,+6H20 >C6H1206+602 Da questa reazione si deduce che sei molecole di CO, devono essere ridotte per ottenere una molecola di glucosio. In realtà la riduzione della CO, avviene interessando una molecola per volta e quindi per ottenere il glucosio è necessario che il ciclo di Calvin si ripeta sei volte; per comodità comunque è meglio descrivere cosa succede ad una singola molecola di anidride carbonica. Entro lo stroma del cloroplasto agisce tutta una serie di enzimi interessa-ti alla fase oscura; il primo di questi si occupa di legare chimicamente la molecola di CO2 con una molecola di un composto normalmente presente in grandi quantità: il ribulosio-1-5-difosfato. L’aggiunta della CO2 rende la molecola molto instabile e dà il via ad una serie di trasformazioni che originano molecole più stabili dal punto di vista chimico. Dapprima la molecola si scinde in due più piccole a tre atomi di carbonio, il 3-fosfoglicerato, le quali vengono poi ridotte con consumo di ATP, utilizzando il NADPH proveniente dalla fase luminosa. Si originano così due molecole di 3-fosfo-gliceraldeide (3-PGA), un composto che può subire diversi destini, compreso quello di servire per la sintesi del glucosio. Bisogna tener presente che i prodotti terminali della fotosintesi non sono soltanto i carboidrati, come si pensava in passato e come si deduce dalla formula generale del processo, ma essi includono anche grassi ed aminoacidi, oltre ad altri tipi di composti organici, che possono tutti essere ottenuti da semplice modificazione enzimatica della 3-fosfo-gliceraldei- de. Durante la normale attività fotosintetica comunque, la maggior parte della GAL-3-P viene trasformata in glucosio, dato che esso rappresenta la miglior molecola in grado di essere accumulata senza grossi problemi. Si è accennato all’inizio della descrizione della fase oscura, che per sintetizzare una molecola di glucosio sono necessarie 6 molecole di CO2, ciò equivale a dire che si formeranno 12 molecole di GAL-3-P, Prova6.9. Solo due di esse sono utilizzate per formare il glucosio, le altre 10 verranno invece ricombinate chimicamente per ricostituire le 6 molecole di ribulosio-1-5-difosfato che hanno dato inizio al processo, tramite il consumo di 6 molecole di ATP: in tal modo il ciclo viene chiuso. La descrizione qui fatta del ciclo di Calvin in realtà è estremamente semplificata, infatti esso comporta almeno dodici reazioni chimiche ben distinte, ognuna catalizzata da un ben preciso enzima. Riassumendo, per ogni molecola di CO2 che entra nel ciclo sono necessari 3 ATP (1 per ripristinare il ribulosio-1-5-difosfato e uno per ciascuna delle due molecole di 3-PGA) e due NADPH (uno per ciascuna molecola di 3-PGA), per cui complessivamente per produrre una molecola di glucosio sono necessari 18 ATP e 12 NADPH. La reazione complessiva della fase oscura sarà quindi: Il glucosio viene immagazzinato entro il cloroplasto e si forma così l’amido primario. Durante la notte la reazione oscura termina e l’amido primario viene pian piano demolito e trasferito attraverso i sistemi conduttori della pianta nelle parti in cui verrà utilizzato, oppure conservato in maniera più duratura, come ad esempio nelle radici. II ciclo di Calvin viene anche chiamato ciclo C3 si vedrà nel capitolo riguardante la foglia che alcune piante hanno evoluto degli adattamenti fotosintetici del tutto particolari, per cui si parlerà anche del ciclo C4 e del metabolismo CAM. |
La Fotosintesi
4568
Messaggio precedenteL'acquario di Simone Balasso
Next PostGioie e dolori nell'allevamento degli Hemichromis





Non ho capito niente 🤔