































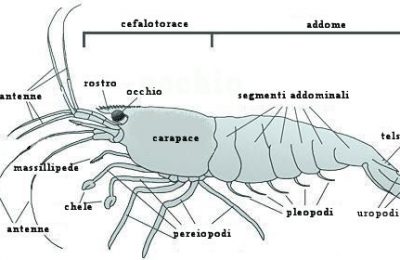















































































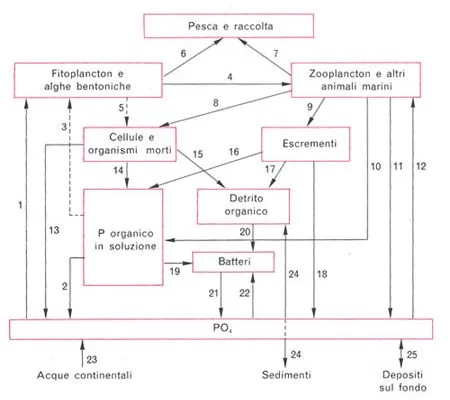
Questo articolo è la stesura di un lavoro svolto in ambito universitario nella materia “Ecologia degli ambienti costieri”. 1) Definizioni di Ecologia, Biocenosi e Biotopo 1) Definizioni di Ecologia, Biocenosi e Biotopo Spesso si tende a privilegiare nella propria visione un aspetto o un altro. Mi spiego: noi acquariofili siamo per lo più affascinati dalla componente vivente quindi ci documentiamo molto bene sulla biocenosi e sui rapporti che intercorrono tra i suoi componenti (gli organismi), spesso tralasciando i rapporti degli organismi con l’ambiente e delle varie componenti del biotopo tra loro. Anche se questo può essere ritenuto per certi versi normale non dobbiamo dimenticare che gli organismi in tutta la loro vita modificano in continuazione l’ambiente (per esempio con l’escrezione o con la respirazione) e subiscono le variazioni ambientali cambiando il loro modo di vivere di conseguenza (entro certi limiti). 2) Vari tipi di rapporti tra i componenti della Biocenosi I rapporti intraspecifici sono di solito legati alla riproduzione, quindi avremo, per esempio, attrazione tra individui di sesso opposto e competizione (che si manifesta mediante comportamenti territoriali e/o lotte più o meno ritualizzate) tra individui dello stesso sesso. Vi sono poi altri rapporti intraspecifici che non rientrano nel novero di quelli legati alla riproduzione: per esempio il cannibalismo (che spesso viene attuato da parte di individui adulti nei confronti di individui giovani) o i comportamenti di branco. Pure i rapporti interspecifici sono vari e interessanti. Uno di questi è la simbiosi. Il più classico esempio di simbiosi è dato dall’accoppiata pesce pagliaccio-attinia, tuttavia ve ne sottoporrò un altro molto più stretto e più importante nella moderna acquariofilia marina: la simbiosi che si instaura tra alcuni invertebrati e le zooxantelle. La maggior parte degli invertebrati che vengono allevati nell’acquario di barriera sono simbionti con zooxantelle e anzi spesso traggono da queste l’unica fonte di nutrimento: ricordiamo gli Actinodiscidi, molti Alcionacei, le Acropora, le Tridacna e così via. Ma cosa sono queste zooxantelle e in cosa consiste il rapporto di simbiosi? Le zooxantelle sono organismi unicellulari e più precisamente Protozoi (animali unicellulari) della Classe Flagellati e dell’Ordine Dinoflagellati. Pur essendo classificati come animali, essi compiono la fotosintesi e forniscono all’invertebrato i polisaccaridi (prodotto della fotosintesi) di cui questo si nutre e si ritiene che negli invertebrati calcificanti (come le Acropora) la simbiosi favorisca la calcificazione. In cambio ne ricavano una protezione da parte di certi loro predatori che vivono nell’acqua aperta. Vi sono poi altri tipi di rapporti interspecifici come l’epibiosi (che si ha nel momento in cui un organismo vive sul corpo di un altro organismo), la parassitosi, e così via. 3) I rapporti di tipo trofico e la divisione tra produttori e consumatori Come si può intuire dall’etimologia dei termini, gli organismi autotrofi sono quelli che riescono a nutrirsi “da soli”, gli organismi eterotrofi sono quelli che per vivere si devono nutrire di altri organismi. Gli organismi autotrofi sono quegli organismi che prendono della materia dal mondo inorganico e la passano al mondo organico. Visto che il passaggio dal mondo inorganico al mondo organico rappresenta un momento in cui molecole più semplici vengono “raggruppate” e “ordinate” a formare molecole più complesse, esso è sfavorito giacché l’universo tende spontaneamente verso il caos e il disordine (più correttamente si direbbe che si tratta di reazioni entropicamente sfavorite). Per ovviare a questo problema gli organismi autotrofi usano una fonte di energia esterna. Pur essendovene altri (certi batteri) gli organismi autotrofi più importanti nella biosfera sono certamente i vegetali o comunque gli organismi che attuano la fotosintesi. Essi prendono dall’ambiente circostante anidride carbonica e acqua formandone molecole organiche, in linea di massima (vi sono altre vie metaboliche collaterali) glucosio, utilizzando come fonte di energia la luce solare (più specificamente le radiazioni nel campo del visibile). Gli organismi eterotrofi (anche l’uomo lo è), invece, non essendo capaci di fotosintesi (né di qualsiasi altro sistema che garantisca l’autotrofia) per vivere sono costretti a cibarsi di altri organismi, scindendoli in molecole più semplici e usando queste per formare le proprie strutture e per le proprie attività metaboliche. Giacché nell’universo nulla si crea e nulla si distrugge gli organismi sono semplicemente dei “trasformatori” di materia, tuttavia gli organismi autotrofi vengono detti “produttori” in quanto a partire da materia inorganica “producono” materia organica, mentre gli organismi eterotrofi sono detti “consumatori” in quanto “consumano” la materia organica “prodotta” dai produttori. Questo “consumo” può avvenire direttamente o indirettamente. Nel primo caso si parla di consumatori di primo grado (come gli erbivori) nel secondo caso si parlerà di consumatori di grado successivo. Per esempio uno Zebrasoma (che si nutre di alghe) sarà un consumatore primario, uno Pterois (che notoriamente è un pesce predatore) nel momento in cui si nutre di un pesce erbivoro si comporta da consumatore di secondo grado e così via. E’ stato notato che il prelievo predatorio (o comunque alimentare) di biomassa (cioè di materia vivente) da un livello trofico è, nei sistemi naturali, non superiore al 10% (principio del decimo ecologico). Nel caso in cui tale prelievo sia superiore o inferiore al 10% il sistema naturale (che ha la caratteristica di essere un sistema autoregolante) perviene ad una nuova situazione di equilibrio che risponda al principio del decimo ecologico. Visto che i consumatori primari possono nutrirsi complessivamente del 10% della biomassa dei produttori, che i consumatori secondari si possono nutrire solo del 10% dei consumatori primari e così via, è chiaro che si verrà a creare una struttura piramidale (piramide trofica o alimentare) a gradini in cui ogni gradino è grande (ovvero ha biomassa) un decimo di quello sottostante (ecco il motivo per cui i predatori apicali, come l’aquila, sono poco diffusi e hanno areali di caccia molto ampi). Il modello della piramide trofica è valido solo nel momento in cui si considerino le categorie dei produttori e consumatori di vario ordine, quando si passa all’esame specifico delle singole specie di organismi il discorso si fa più complesso. Infatti molti organismi si comportano da consumatori non di un determinato ordine. Pensiamo a noi uomini: ci nutriamo indifferentemente di vegetali (comportandoci da consumatori di primo grado), polli e tacchini (che sono erbivori, e quindi ci comportiamo da consumatori di secondo grado), pesci (che potrebbero essere consumatori anche di terzo grado). Oppure, tornando all’esempio di prima, allo Pterois non interessa se il pesciolino che gli passa davanti è erbivoro o si nutre di piccoli animali, se lo mangia e basta! A complicare il quadro vi sono poi gli organismi detritivori e i batteri. Infatti i detritivori, come certi policheti erranti comuni negli acquari di barriera detti dagli acquariofili “vermi setolosi” (bristleworms) o “fireworms” (per via del fatto che sono urticanti), si cibano di sostanza organica detritale (cioè persa dai livelli trofici non in seguito a predazione). Per esempio un organismo che muore scende al fondo diventando, dopo un certo tempo, sostanza organica detritale (o più semplicemente detrito). I detritivori oltre al detrito ingeriscono poi anche i batteri saprofiti su di esso. Per quanto riguarda i batteri, poi, la loro collocazione nel quadro della piramide alimentare è, se possibile, ancora più complessa (ricordiamo che vi sono batteri organicatori di varie sostanze). In base a quanto detto, quando si passa ad un esame specifico dei rapporti tra gli organismi di una data biocenosi si preferisce sostituire al concetto di piramide trofica quello di rete trofica. Tra i vari punti della rete trofica (i vari organismi) avviene uno scambio, più o meno diretto, di materia ed energia (in quanto gli atomi delle molecole sono tenuti insieme da legami energetici). 4) Cicli di energia e materia Per quanto riguarda la materia, l’uomo ha sentito il bisogno di dividere il ciclo della materia nei sistemi naturali nei cicli dei vari elementi, si parlerà quindi di ciclo del Carbonio, ciclo del Fosforo e così via. Qui se ne esamineranno i principali con riferimento all’ambiente marino. Ciclo del Carbonio: il Carbonio (C) entra nell’ambiente marino o attraverso le immissioni fluviali di carbonio detritale (generalmente costituito da vegetali morti), tuttavia spesso questo carbonio detritale è refrattario ad entrare nel ciclo, oppure vi entra sotto forma di anidride carbonica atmosferica che passa in soluzione secondo questo schema di reazioni: CO2 + H20 <=> H2CO3 <=> H+ + HCO3- <=> 2H+ + CO32- Spieghiamo la formula su esposta: l’anidride carbonica (CO2) si combina con l’acqua (H20) a formare acido carbonico (H2CO3 ) il quale, tuttavia, in acqua è in larga parte dissociato in un protone (ione H+) e nello ione HCO3- , il quale in parte tende a perdere un altro protone a formare lo ione carbonato (CO32-). Lo ione carbonato è molto importante nell’ambiente marino e pure per l’acquariofilo di barriera. Infatti gli organismi costruttori delle barriere coralline hanno uno scheletro costituito da carbonato di calcio (CaCO3) e alcuni di essi sono interessanti ospiti dei nostri acquari. Il carbonato di calcio si può trovare in tre possibili diverse forme (dette forme allotropiche): calcite (come quella delle valve delle conchiglie), aragonite (lo scheletro della maggior parte dei coralli è appunto costituito da aragonite) e vaterite (poco diffusa). I cicli dell’Azoto e del Fosforo: i composti dell’Azoto (N) e del Fosforo (P) sono detti nutrienti in quanto, essendovi nelle acque marine superficiali abbondanza di luce, acqua e anidride carbonica, costituiscono l’unico fattore limitante per lo sviluppo degli organismi fotosintetici. Le concentrazioni di questi elementi nelle acque è molto variabile (ad esempio la concentrazione di nitrati varia circa da 0,02 a 30 mg/l). Nelle acque con grosse quantità di nutrienti (come nelle zone in cui vi sono scarichi industriali o urbani, o presso le foci dei fiumi) si assiste ad un fenomeno detto “eutrofizzazione delle acque” che consiste nello sviluppo massiccio di organismi fotosintetici unicellulari ad alta velocità riproduttiva (di solito Dinoflagellati) che colorano le acque. In ambienti con scarsa quantità di nutrienti è necessaria una forte specializzazione degli organismi che devono anche cercare di abbassare la competizione interspecifica per l’utilizzo delle risorse: ecco comparire l’immensa varietà di forme viventi e associazioni (come la già menzionata simbiosi tra invertebrati e zooxantelle) che caratterizza la vita delle barriere coralline, ambienti che sono notoriamente poveri di nutrienti. L’entrata di azoto nell’ambiente marino può avvenire o per mezzo dei già menzionati afflussi fluviali o antropici o, ancora, per fissazione dell’azoto gassoso atmosferico (N2) da parte di particolari ceppi batterici (detti per questo “batteri fissatori dell’azoto”). L’azoto fissato dai batteri passa ai livelli trofici superiori a mezzo degli organismi batterivori, ma in parte torna in soluzione per morte o escrezione. Questo azoto passa dalla forma NH4+ (ione ammonio) alla forma NO2- (ione nitrito) ad opera dei batteri del genere Nitrosomonas, i batteri del genere Nitrobacter trasformano gli ioni nitrito in ioni nitrato (NO3-).
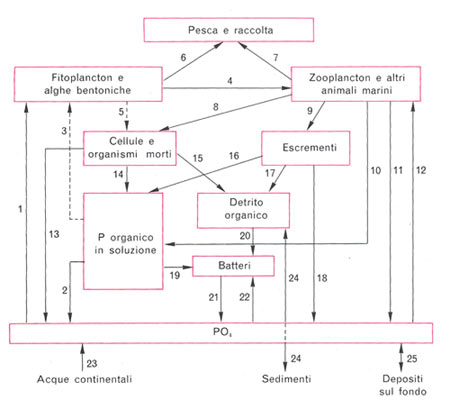
Questi processi avvengono in ambiente aerobio (cioè in presenza di ossigeno) ed è appunto su questi processi e su questi ceppi batterici che si basa l’attività del filtro biologico tradizionale il quale non è altro che una zona ove si favorisce l’insediamento di una grossa quantità di questi batteri aerobi. I nitrati vengono assorbiti dai vegetali che li utilizzano e li passano ad altri livelli trofici (venendo mangiate dagli erbivori). Ovviamente anche in questi punti del ciclo si ha perdita di azoto per morte o escrezione. L’uscita di azoto dall’ambiente marino (a parte quella che si realizza per esempio mediante la predazione di pesci da parte degli uccelli marini) avviene ad opera di altri batteri. Questi batteri, che vivono in ambiente anaerobio (cioè in assenza o scarsità di ossigeno, nella zona più profonda delle rocce e dei sedimenti), trasformano i nitrati in azoto gassoso il quale, in superficie, passa all’atmosfera. E’ su questi ceppi batterici che si basano il filtro denitratore e l’uso di pietre vive nella moderna acquariofilia di barriera. Per quanto riguarda il ciclo del fosforo, esso entra nell’ambiente marino mediante apporti di acque fluviali che avevano dilavato rocce contenenti fosfati (PO43-). I fosfati vengono utilizzati dai vegetali che li passano ai livelli trofici superiori. Ogni livello, al solito, perde parte dei fosfati per morte o escrezione (una parte di questi fosfati può anche sedimentare). La fuoriuscita dall’ambiente marino dei fosfati avviene principalmente ad opera degli uccelli marini i quali nutrendosi di pesce sottraggono i fosfati dall’acqua e defecando presso le scogliere causano accumuli di guano che nel corso del tempo vanno a costituire le rocce fosforitiche (quelle che prima abbiamo visto essere dilavate dalle acque fluviali), chiudendo così il ciclo.
Ciclo dello Zolfo: il ciclo dello Zolfo (S) prevede un’entrata di zolfo nell’ambiente marino ad opera dei fiumi e delle piogge che immettono nell’ambiente marino solfati (ioni SO42-). I solfati vengono utilizzati da batteri e vegetali che passano così lo zolfo ai livelli trofici superiori dai quali sfuggono per morte o escrezione, passando nel sedimento. Nella zona meno superficiale del sedimento, in ambiente anaerobio, i solfati vengono ridotti ad acido solfidrico dai ceppi batterici Desulfovibrio e Desulfomatuculum (detti tiobatteri). L’acido solfidrico, reagendo con altre sostanze, può formare composti che non torneranno in soluzione oppure, passando in zone ricche di ossigeno del sedimento, essere di nuovo trasformato in solfati, sempre ad opera di batteri. 5) Strategia R e strategia K In sostanza la strategia R prevede una forte adattabilità alle più svariate condizioni ambientali con utilizzo di un ampio range di risorse, cosa che, per contro, implica una scarsa specializzazione nell’utilizzo di tali risorse e, quindi, un loro utilizzo non ottimale. La strategia R prevede che la riproduzione porti ad un alto numero di individui in poco tempo di modo che in mezzo al gran numero di individui prodotti, una piccola parte sopravviva. Le specie che presentano fondamentalmente una strategia R sono dette “specie pioniere” o “specie fuggitive” dato che, nelle successioni di popolamento a partire da un ambiente senza nessuna specie, sono le prime a comparire e ad invadere l’ambiente utilizzando le risorse (pensiamo alle erbacce nei campi abbandonati o alle alghe patinose che si sviluppano pochi giorni dopo l’allestimento di un acquario marino). Esse, poi, vengono col tempo soppiantate da specie meglio adattate alle condizioni ambientali, che utilizzano un range limitato di risorse, ma lo utilizzano al meglio. Queste ultime sono le specie a strategia K, caratterizzate, dal punto di vista dell’attività riproduttiva, dalla produzione di un basso numero di individui ma dotati di grossa aspettativa di sopravvivenza. Come detto, le popolazioni di queste specie compaiono solo in un secondo momento nelle successioni di popolamento lasciando, dopo un certo periodo di tempo, le specie pioniere negli ambienti di margine, sottoposti a maggiori escursioni dei parametri ambientali. Il discorso su queste strategie riveste una notevole importanza anche per l’acquariofilo in quanto, in linea di massima, una specie a spiccata strategia K sarà molto difficile (se non impossibile) da allevare in acquario in quanto adattata a caratteristiche ambientali altamente specifiche e che praticamente non variano mai; viceversa una specie a spiccata strategia R sarà tanto facile da allevare in acquario (anche se a noi potrebbe non fare piacere) da risultare infestante, specie in vasche con caratteristiche instabili. Prendiamo per esempio un Gorgoniaceo: non sarà certo un organismo facile da allevare, perché adattato a vivere nell’acqua profonda ove le caratteristiche ambientali variano pochissimo. Prendiamo poi un rametto di Caulerpa: se non stiamo attenti potrebbe anche risultare infestante in quanto utilizza un’ampia gamma di risorse. Prendiamo, infine, un acquario con caratteristiche fortemente instabili: non ci dovremmo preoccupare di cercare certi organismi (come certe alghe patinose) saranno loro a cercare noi! 6) Biomassa e produzione La biomassa è costituita dalla massa vivente. Benché vi siano ambienti particolari con ampi substrati da colonizzare e poca acqua (come le pozze di scogliera) in cui i produttori più importanti sono le macroalghe o altri ambienti in cui lo sono le fanerogame marine (come la Posidonia), i più importanti produttori, considerato il mare nel complesso, sono organismi fotosintetici microscopici, generalmente unicellulari che vengono spesso genericamente raggruppati sotto il termine “fitoplancton”. La misurazione della biomassa fitoplanctonica può avvenire in diversi modi, ognuno dei quali ha dei difetti: misurazione del contenuto di ATP o del contenuto di clorofilla di un dato campione (tale sistema ha lo svantaggio che tali sostanze variano nel corpo degli organismi in base alla loro attività metabolica), misurazione del contenuto di DNA del campione (insieme al DNA vivente viene misurato il DNA detritale che può essere presente pure in misura rilevante). La variazione della biomassa dei produttori nell’unità di tempo è detta produttività primaria (da alcuni impropriamente detta produzione primaria) ed è importante in quanto da essa dipendono i livelli trofici superiori. La produttività primaria fitoplanctonica viene misurata in laboratorio ponendo in date condizioni (luce, nutrienti) una coltura di fitoplancton e controllando la quantità di carbonio marcato (14C, fornito mediante bicarbonato di sodio marcato) che viene fissata nell’unità di tempo. La produttività primaria, com’è ovvio, viene influenzata da quei fattori che influenzano la vita dei vegetali quindi, nell’ambiente marino, la luce (nelle zone superficiali, che sono quelle di maggior interesse acquaristico, generalmente presente in gran quantità), la presenza di nutrienti (che varia in base a una notevole quantità di fattori come le correnti sottomarine, l’apporto di acque fluviali, l’attività antropica e così via), la temperatura (in quanto, entro certi limiti, più alta è la temperatura più rapido è il metabolismo degli organismi). La variazione della biomassa dei consumatori nell’unità di tempo è detta produttività secondaria e varia in base alla temperatura e alla disponibilità e alla quantità di alimento. Concludo augurandomi di aver fornito alcune nozioni di ecologia dell’ambiente marino utili all’acquariofilo per comprendere meglio alcuni fenomeni che accadono al di là del vetro del loro acquario. Bibliografia: -M. La Greca-ZOOLOGIA DEGLI INVERTEBRATI-UTET -N. Della Croce, R. Cattaneo Vietti, R. Danovaro -ECOLOGIA E PROTEZIONE DELL’AMBIENTE MARINO COSTIERO-UTET
|
Cenni di Ecologia per l’acquariofilo marino
2663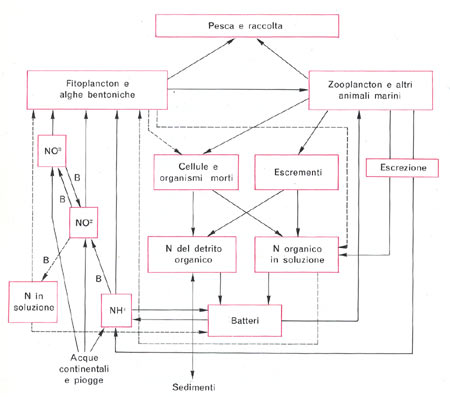
Messaggio precedenteRealizzazione di un acquario di barriera di 180 litri
Next PostL' acquario di Marco Serazio



