














































































































In questo articolo forniremo un’analisi dettagliata delle strutture anatomiche che caratterizzano questi affascinanti organismi acquatici. Scoprirete gli intricati dettagli delle pinne, delle scaglie, degli organi interni e dei sistemi sensoriali che consentono ai pesci di adattarsi e prosperare nell’ambiente acquatico.
Ci concentreremo su aspetti come la locomozione, la respirazione, la digestione e la percezione sensoriale.
Alla fine di questa lettura avrete una comprensione chiara e completa di questi animali.
Squame dei pesci
Le squame o scaglie cutanee sono formazioni dermiche situate in strati profondi dell’epidermide.
Tali scaglie proteggono efficacemente gli animali dalle ferite; nel corso dell’evoluzione sono comparsi vari tipi di scaglie.

Una squama vista al microscopio
Quelle denominate placoidi, degli squali e delle specie affini del gruppo dei Condritti sono, effettivamente, denti epidermici, con una superficie di smalto che ricopre una polpa di dentina; nella cavità boccale, grandi scaglie placoidi formano file di denti.
Si registrano altri diversi tipi di squame: cicloidi, ctenoidi e ganoidi (le tipologie riguardanti i pesci d’acqua dolce).





Vari tipi di squame.
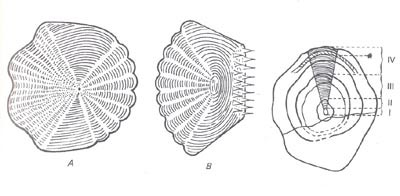
Le scaglie cicloidi sono alquanto diffuse e di origine primitiva e sono altresì dette per la loro foggia tondeggiante, caratterizzata da un margine liscio; sono presenti nei ciprinoformi.
Le scaglie ctenoidi sono connotate da una superficie ruvida e un margine munito di dentelli, la cui forma ricorda quella dei denti di un pettine; sono rintracciabili nei perciformi.
Le 2 tipologie summenzionate palesano sulla loro superficie un intreccio di formazioni concentriche che rappresentano uno strumento imprescindibile per stabilire l’età dell’animale.
Le scaglie ganoidi rappresentano una tipologia primitiva suddivisa in 2 rami principali: il paleoniscoide attinente all’estinto Paleoniscus, ma esistente anche in specie attuali; il secondo riguarda una tipologia evoluta denominata lepisosteoide.
Tali scaglie sono connotate spesso da una forma romboidale e sono strutturate tra loro in modo parzialmente sovrapposto, e si trovano nei politteri, gli storioni e le amie.
Lo strato superiore di queste scaglie è ricoperto da uno strato di ganoina che dà loro un aspetto vitreo e brillante e che ha la stessa origine della dentina.
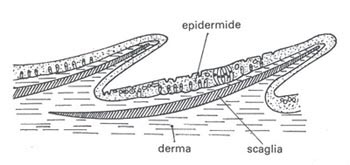
Le scaglie dei pesci ossei (teleostei) hanno la forma di sottili placchette ovali, sprovviste di smalto e di dentina.
Le scaglie cosmoidi sono una modificazione delle ganoidi, ed assomigliano alle scaglie placoidi, che nei pesci attuali si trovano soltanto nel celacanto.
(da L.Bertin)
Come sono fatte le ossa dei pesci
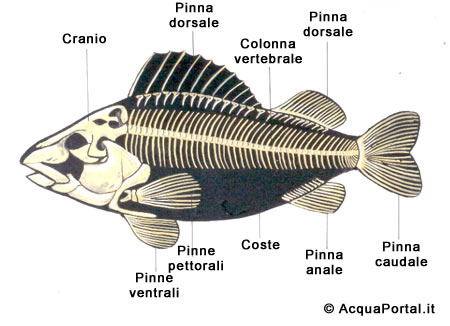
Lo scheletro consta di 2 differenti parti: lo scheletro appendicolare comprende le parti scheletriche delle pinne e i cinti pettorale e pelvico; lo scheletro assiale raggruppa la scatola cranica e la colonna vertebrale.
Il neurocranio è costituito da un’architettura scheletrica piuttosto complessa che si basa su blocchi compatti di ossa di vario tipo denominate autostosi (otiche, temporali, mesetmoide, sfenoidale) e allostosi (setti nasali, il vomere, i setti frontali, parietali, frontali, orbitali, lacrimali, iugali, squamosi).
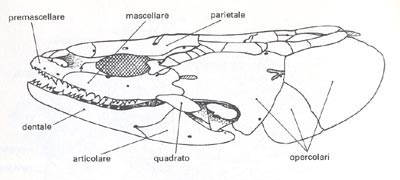
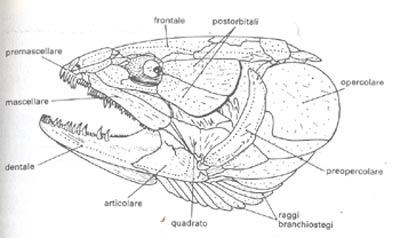
Altra parte che concorre alla formazione del cranio è lo splancnocranio, composto da una grande quantità di ossa di tipo mobile, di origine opercolare che rivestono le fessure branchiali, e relative alle arcate viscerali.
Il cranio degli squali, delle razze e delle chimere è cartilaginoso, tuttavia, in alcune specie sono presenti incrostrazioni secondarie di sali di calcio, che gli conferiscono un aspetto osseo.
Gli storioni presentano un cranio cartilaginoso, ma vengono catalogati con quelli ossei, poichè i loro antenati avevano lo scheletro ossificato.
Caratteristiche le forme ossee denominate premascellari, che dotate di una certa mobilità, permettono alla bocca di sporgersi esternamente; completano la formazione della mascella superiore le ossa mascellari (esse possono anche fungere da supporto ad una struttura dentale).
Ma le lamprede, analogamente alle missine, non presentano le mascelle, in quanto rimpiazzate da una ventosa boccale; gli archi branchiali sono sostenuti da supporti cartilaginosi autonomi.
La colonna vertebrale è globalmente ossificata composta da vertebre a mo’ d’arco, sviluppatesi dalle basi di archi neurali e adiacenti nella parte caudale ad archi ematici, che si possono descrivere come “portali” in cui si snodano arteria e vena caudali.
Nelle lamprede e nelle missine, si tratta ancora di una corda dorsale sprovvista di tessuto cartilaginoso od osseo.
Niente annuncia ancora le vertebre, e la coda è semplicemente sormontata da piccole formazioni cartilaginose.
Nei pesci cartilaginei, le vertebre sono evidenti; sono cartilaginose ed avvolgono la corda interna; sono uniti mediante apofisi.


Sulla parte dorsale si trova un canale che contiene il midollo spinale; nella cavità generale, le costole si attaccano sul lato ventrale delle vertebre.
Nell’area caudale la base delle vertebre comprende un canale che protegge la vena e l’arteria caudale.
La maggior parte dei pesci ossei ha le vertebre conformate allo stesso modo, ma ossificate, tuttavia vi sono delle eccezioni, come nel caso dello storione.
Le coste sono elementi ravvisabili nelle vertebre, che hanno la funzione di separare le viscere dalla massa muscolare.
In alcuni generi è presente una costa di tipo dorsale che svolge una funzione analoga, ma tra parti differenti di origine muscolare, nella fattispecie dorsale e ventrale.
Lo scheletro delle pinne è impari, ed è formato da raggi ossei e cartilaginose; le pinne pettorali dei pesci ossei (teleostei) sono saldate al cranio, mentre negli animali cartilaginei, sono collegate con lo scheletro dell’apparato branchiale. In ambedue i gruppi, le pinne pelviche sono fissate liberamente nella muscolatura, con l’eccezione di alcuni gruppi, quali i gadiformi: le loro pinne ventrali sul davanti dei pettorali e sono collegate al cranio.
I denti dei pessci
La dentizione anteriore è composta dai Vomerini, situati nella parte alta della bocca, Palatini, posti lateralmente, e i Premascellari.
La dentatura posteriore è caratterizzata da denti faringei derivanti da formazioni delle arcate branchiali.

di un Piranha
Grazie a questi particolari denti, i Ciprinidi, ad esempio, possono sopperire alla carenza di denti veri e propri.
Tali denti possono assumere diverse forme a seconda del genere; segnaliamo la caratteristica forma a cucchiaio di B. brachycephalus caspius, il dente da macina della carpa, e il dente simil canino, di Aspius aspius, utilizzato principalmente per catturare le prede, i denti lunghi ed acuminati di Bathybates leo o Belonesox belizanus.
Le pinne
Si registrano 2 tipologie di pinne: pinne pari e pinne impari.
Le prime sono sorrette da un supporto scheletrico basato su determinate strutture a forma d’arco (cinto pettorale e cinto pelvico), e comprendono le pinne pettorali e pelviche.
Tali pinne sono sorrette da raggi cutanei (lepidotrichi) appartenenti alla membrana dell’arto, e da raggi endoscheletrici profondi.
I raggi lepidotrichi sono posizionati su ciascun lato della pinna e sono caratterizzati da un rivestimento sotto pelle di piccole lamine ossee separate tra loro, assimilabili a scaglie cutanee, che conferiscono ai raggi una flessibilità in corrispondenza dei lati.
In virtù di queste caratteristiche, tali raggi cutanei sono molli; qualora, però, le placche ossee si plasmino in un corpo unico, il raggio assume la foggia di un aculeo, che può costituire una potente arma per l’animale che ne fa uso.
Il cinto pettorale (o scapolare) è rimarcato da un processo di ossificazione più o meno evidente, ed è formato dalla scapola, posizionata nella regione dorsale, il caracoide, presente in corrispondenza della zona ventrale, che in posizione laterale entrano in contatto tra loro e con una parte dell’arto.
Tramite la clavicola e il cleitro il cinto scapolare è collegato alla parte posteriore del neurocranio.
Il cinto pelvico, invece, presenta uno sviluppo sensibilmente minore, contrassegnato da un supporto scheletrico compatto che però non crea alcun collegamento tra le pinne e la colonna vertebrale.
Le pinne pari consentono lo spostamento dei pesci nell’ambiente acquatico; nei pesci ossei, le pettorali sono collegate al cranio, mentre negli animali cartilaginei sono ancorate nella muscolatura mediante elementi cartilaginosi autonomi.
In genere, le pinne pelviche sono saldate, analogamente alle pettorali dei pesci pesci cartilaginei, nella muscolatura.

In alcuni gruppi, come i clupeiformi ed i ciprinoformi, le pinne pelviche si trovano in posizione ventrale o addominale; nei preiformi sono situate sotto le pettorali e nei gadiformi sono posizionate proprio davanti ai pettorali, tuttavia in alcuni gruppi sono del tutto assenti, come nel caso delle anguille.
Le pinne impari includono uno o più arti dorsali, una pinna caudale e una o più anali, e sono composte da raggi endoscheletrici mobili che si biforcano tra loro.
Tali raggi sono confinanti con le emospine della colonna vertebrale, per questo motivo è aleatorio stabilire se tali raggi siano originati da pezzi vertebrali, o rappresentino uno sviluppo a se stante.
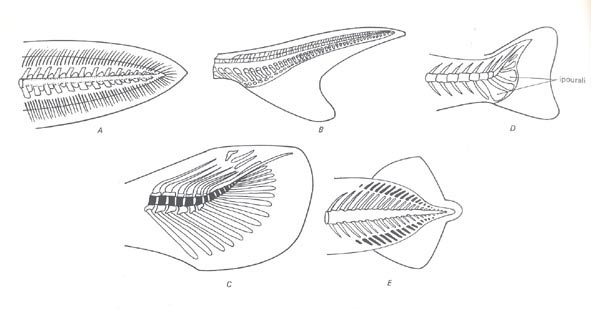
La pinna caudale è omocerca, di forma simmetrica (ma asimmetrica internamente), poichè caratterizzata da una ripartizione in 2 parti uguali, nella fattispecie lobo dorsale e ventrale.

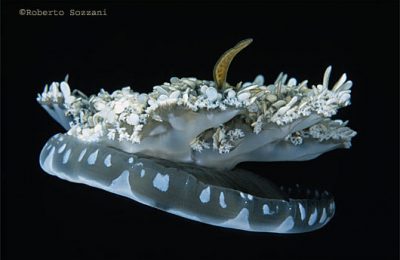
(Foto Roberto Sozzani).
La fusione delle 2 parti succitate determina la formazione dell’urostilo, in cui sono evidenti emospine modificate denominate epiurali.
Si registra la presenza di pinne caudali di forma eterocerca (con le 2 parti summenzionate di forma asimmetrica), dificerca (arti eterocerchi trasformatisi in un secondo tempo in pinne omocerche, anche per quanto concerne lo scheletro interno), ed emiomocerca.
Ad esclusione delle pinna di posa, tutte le altre pinne sono rese solide dai raggi ossei o cartilaginosi, e possono modificarsi in organi copulatori che permettono una fecondazione interna (ad esempio, le pinne ventrali possono trasformarsi in mixopterigi o pterigiopodi negli squali, oppure in ventose, come nel caso delle pinne ventrali dei gobidi).
Motilità
In genere, i pesci nuotano mediante ondulazioni di tutto il corpo (anguille), o soltanto il peduncolo caudale (come avviene per la maggioranza delle specie).
Alcuni pesci compiono anche movimenti ondulatori delle pinne pari (pesci cofano, notopteridi, ecc).
Anche i movimenti sincroni o asincroni delle pinne pari producono risultati ragguardevoli: alcune specie della famiglia Gasteropelecidae sono addirittura in grado di uscire dall’acqua, e fanno dei voli accompagnati dal battito attivo delle pinne pettorali.
Al contrario, il volo dei pesci marini di determinate specie (Exocoetidae e Hemirhampidae) è assolutamente passivo: le loro pinne pettorali, che arrivano all’80% della lunghezza del corpo consentono loro di planare nell’aria, effettuando spostamenti che possono superare i 400 m, quando la forza del vento è propizia.
La forma del corpo e la sua superficie liscia sono fondamentali per ridurre la resistenza dell’acqua; di non minor importanza anche la morfologia della pinna caudale.
Nei nuotatori provetti, i lobi superano la zona delle correnti con le reazioni turbolente che si formano sul retro della dimensione massima del corpo e prendono parte attiva all’orientamento del pesce in fase di spostamento.
La velocità massima che il pesce raggiunge nell’arco di brevi periodi di tempo varia da specie a specie: dai 3 m al secondo, coperti dalla lucioperca, fino ai 36 del pesce spada.
Tuttavia, questi valori sono destinati a cambiare se si considera la velocità media raggiunta da ciascuna specie.
La muscolatura
La muscolatura dei pesci non si differenzia granchè da una specie all’altra.
Tra i muscoli si trovano numerose lische sottili, spesso biforcute, non collegate allo scheletro; queste sono caratteristiche dei pesci, e compaiono per ossificazione all’interno di custodie connettive di fibre muscolari isolate.
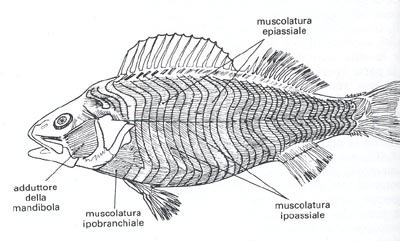
Per trovare muscoli più differenziati occorre esaminare la testa; gli organi muniti di muscoli autonomi sono i bulbi oculari, l’apparato branchiale e le mascelle.
La muscolatura del tronco è concentrata soprattutto nella parte dorsale del corpo, ed il setto orizzontale la divide in 2: muscolatura superiore e muscolatura inferiore.
A loro volta, i muscoli di dividono in scheletrici striati, gestiti attivamente dall’animale, e viscerali lisci, indipendenti dalla volontà; il muscolo cardiaco, per quanto striato, non risponde agli ordini volontari.
Parte della muscolatura scheletrica denota un cromatismo rosso più accesso della rimanente, ed è situata proprio sotto l’epidermide, nella parte mediana del corpo, al livello del setto orizzontale, e nel peduncolo caudale.
La colorazione dei muscoli rossi è dovuta ad un’elevata concentrazione di emoglobina e di mioglobina.
I muscoli rossi possono lavorare prolungatamente senza stancarsi: questa è la ragione per cui le specie che compiono lunghi tragitti presentano una proporzione elevata di tali muscoli.
I muscoli chiari, ossigenati di meno, si stancano presto, ma possono compiere grandi sforzi su percorsi brevi, che si pagano però con una carenza di ossigeno che porta ad un aumento della concentrazione di acido lattico.
E’ soltanto quando quest’ ultimo viene eliminato che i muscoli bianchi sono pronti ad operare nuovamente; tali muscoli costituiscono la maggioranza (pochissimi, invece quelli rossi) nei pesci dotati di scarsa mobilità.
Colori e pigmenti nei pesci
L’epidermide è composta da un rivestimento multistrato, caratterizzato da uno spessore variabile in relazione ad una determinata specie o ad una zona particolare del corpo.
In tali strati epidermici abbondano ghiandole unicellulari e mucipare; queste ultime sembra adempiano alla salvaguardia dell’animale dall’insorgenza o meno di eventuali infezioni di origine batterica, fungicida e altro.
Inoltre, può essere registrata la presenza di ghiandole velenifere, riconducibili a lamette situate alla base delle pinne, oppure di fotosfori, organi luminosi usati dai pesci che vivono nelle zone molto profonde ammantate nell’oscurità o scarsamente illuminate.
Nel derma (la parte sottostante l’epidermide) sono presenti cellule pigmentarie (cromatofori e iridociti) che, in base a determinate condizioni ambientali percepite dal pesce, possono modificare il colore o la decorazione della pelle.
I cromatofori si suddividono in eritrofori, caratterizzati da pigmenti rossi e arancioni, xantofori, muniti di pigmenti gialli e melanofori, la cui pigmentazione risulta nera.
Il regime alimentare da somministrare ai pesci in cattività si rivela importante anche per salvaguardare la vivacità cromatica delle loro livree, infatti alcuni pigmenti sono contenuti nelle sostanze alimentari ingerite dai pesci.
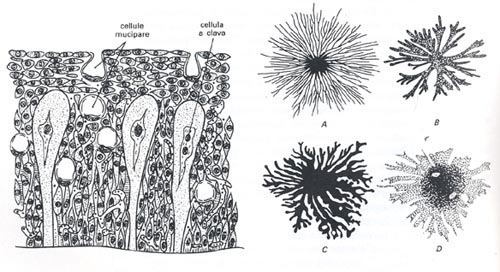
Naturalmente le colorazioni suggestive che ammiriamo in alcune specie sono il frutto della combinazione cromatica dei tipi di cromatofori summenzionati.
Gli iridociti, costituiti da sostanze di guanina (un sottoprodotto della digestione delle proteine), concorrono a trasformare la colorazione base di una livrea in un cromatismo argenteo e iridiscente.
L’animale può modificare la sua colorazione a seconda di sollecitazioni differenti: la necessità di mimetizzarsi nell’ambiente per fronteggiare situazioni pericolose o attendere una preda di passaggio, crea le premesse per tale mutazione; Betta splendens, ad esempio, può attuare tale processo quando è attaccato da un contendente; i cromatismi assumono tonalità differenti durante il periodo riproduttivo.
I pesci modificano la loro colorazione durante il loro sviluppo, nei periodi di migrazione, a seconda delle stagioni, e in base al loro stato umorale e psicologico.
In genere, gli esemplari giovani presentano una colorazione differente da quella degli adulti, e questo corrisponde alla differenza del mezzo in cui vivono.


E’ però durante la fregola che si assiste ai cambiamenti più spettacolari: i maschi assumono colori smaglianti, senza preoccuparsi della loro visibilità, ma tale arricchimento cromatico è ravvisabile soltanto da pozioni molto ravvicinate.
Alcuni pesci possono reagire molto velocemente alla variazioni cromatiche del supporto sul quale si trovano, infatti possono adattarsi al colore predominante dell’ambiente.

(Foto Roberto Sozzani).
Anche i colori premonitori di certi animali che vivono nell’habitat corallino, servono ad avvertire gli altri esemplari che il territorio circostante non può essere violato.
I colori intensi delle specie di scoglio derivano dalla loro vita fra i boschetti di coralli colorati.
La secrezione di determinate sostanze (originarie delle ghiandole endocrine), causata dalla stimolazione ormonale, concorre alla variazione cromatica della livrea.
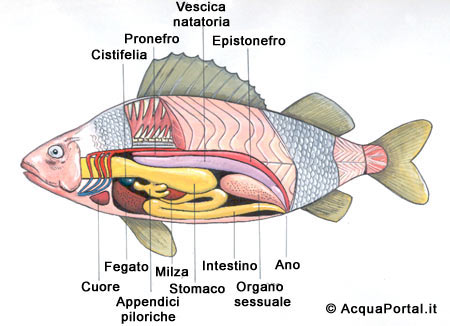
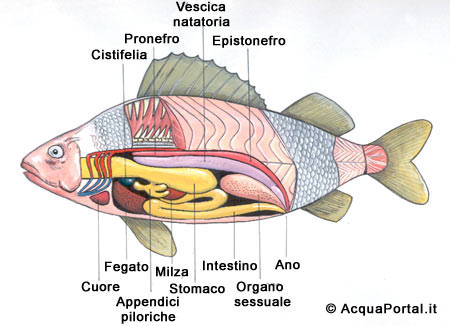
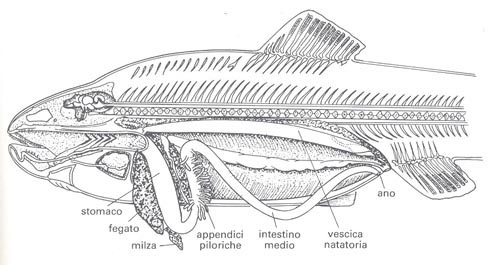
L’apparato digerente
La struttura organica di tale apparato consta di diversi organi, nella fattispecie, faringe, esofago, cavità orale, stomaco ed intestino, a cui sono collegati altri organi che lavorano in sinergia con quelli succitati: denti, lingua, pancreas, fegato e cistifellea.
In bocca non sono presenti ghiandole salivari, ma soltanto ghiandole mucose.
La lingua non è dotata di muscolatura, in quanto costituisce una semplice protuberanza del pavimento boccale frapposta tra le file dentali; la cavità orale non hanno uno sviluppo accentuato poiché termina in corrispondenza della faringe, quindi è piuttosto breve.
E’ importante constatare che la lingua non è particolarmente sviluppata nei pesci ossei, a differenza di quanto avviene per i ciclostomi.
I denti, a seconda della famiglia di appartenenza, possono muoversi o essere saldati solidamente alla struttura ossea sottostante.
L’esofago è piuttosto largo, ma presenta uno sviluppo breve e lineare; lo stomaco è caratterizzato da tratti ascendenti e discendenti che gli conferiscono una forma di sacca.
Le pareti di tale organo sono dilatabili per consentire all’animale (in genere specie predatrici) di ingerire prede piuttosto voluminose.
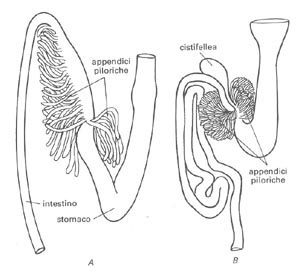
Tuttavia, alcuni generi di pesci (Protopterus e Ciprinus, ad esempio) sono carenti di stomaco, grazie ai loro efficienti apparati masticatori che gli consentono di tagliuzzare abbondantemente il cibo ingerito, e quindi di completare agevolmente il processo digestivo con l’ausilio degli enzimi prodotti dall’intestino.
L’intestino è di forma particolarmente allungata nelle specie fitofaghe, così formato per favorirne l’assimilazione di sostanze vegetali, viceversa negli animali carnivori presenta una morfologia corta.
Nei salmonidi o nei gadaci, tale organo forma un gran numero di diverticoli che accrescono la superficie digerente, e probabilmente contribuiscono anche a neutralizzare la reazione acida del bolo alimentare all’uscita dello stomaco.
B) pesce carnivoro
A differenza degli enzimi digestivi dello stomaco, di natura acida, quelli intestinali richiedono un ambiente lievemente basico.
E’ nell’intestino che vengono digerite le proteine, con l’ausilio della tripsina, mentre altri enzimi (lipasi, carbossilasi) agevolano la digestione di sostanze grasse e degli zuccheri.
La grandezza del fegato è piuttosto rilevante, ed è caratterizzato da un’abbondante concentrazione di grassi; in esso è diffuso il pancreas (importante per la secrezione dell’insulina, necessaria per mantenere la glicemia al suo tasso ottimale), la cui distribuzione cellulare è capillare, in quanto non concentrata in un determinato punto.
Inoltre, il fegato produce la bile, che favorisce la digestione dei lipidi ed aumenta il p H del contenuto intestinale, e conserva lo zucchero del sangue, i suddetti lipidi e le vitamine A e D.

